|
| Fibulacaris nereidis / Artwork by Danielle Dufault @MesozoicMuse |
The origin of the arthropod carapace, an enlargement of cephalic tergites, can be traced back to the Cambrian period. Even so, its disparity and evolution are still not fully understood. It is the detailed study of species such as this new ‘bivalved’ arthropod, Fibulacaris nereidis gen. et sp. nov., that will help us get closer to the truth.
 |
| Interpretive Cladogram |
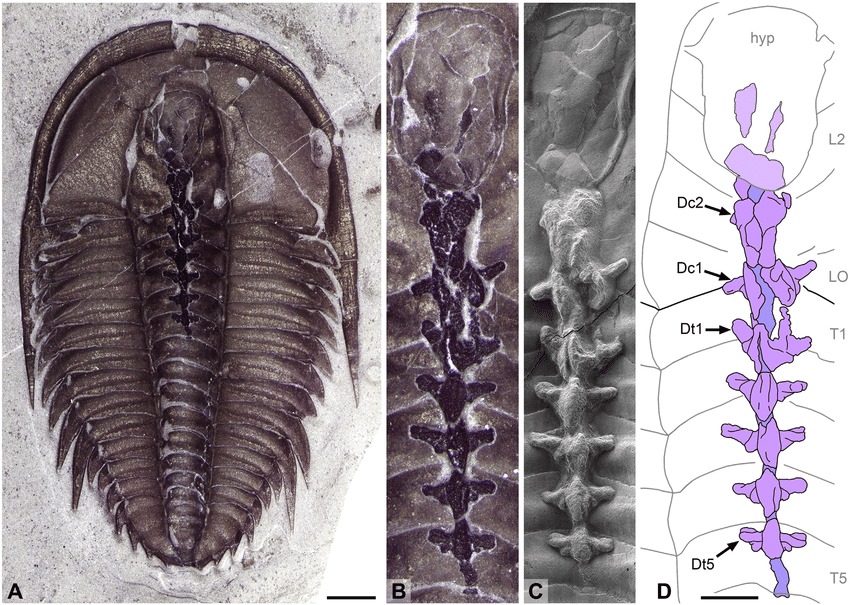
The laterally compressed carapace covers most of the body. It is fused dorsally and merges anteriorly into a conspicuous postero-ventrally recurved rostrum as long as the carapace and positioned between a pair of backwards-facing pedunculate eyes.
The body is homonomous, with approximately 40 weakly sclerotized segments bearing biramous legs with elongate endopods, and ends in a pair of small flap-like caudal rami. Fibulacaris nereidis is interpreted as a suspension feeder possibly swimming inverted, in a potential case of convergence with some branchiopods.
A Bayesian phylogenetic analysis places it within a group closely related to the extinct Hymenocarina. Fibulacaris nereidis is unique in its carapace morphology and overall widens the ecological disparity of Cambrian arthropods and suggests that the evolution of a ‘bivalved' carapace and an upside-down lifestyle may have occurred early in stem-group crustaceans.
Fibulacaris nereidis contributes to the increasing morphological, functional, ecologic and taxonomic diversity of bivalved arthropods known from the Cambrian period. The shape of the carapace, with its single posteriorly directed ventral rostrum, appears to be morphologically unique not only among Cambrian and other fossil species but similarly rare across extant crustaceans or other arthropods. The carapace, including the rostrum, most probably had a protective role, but as in other extant arthropods, could have contributed to swimming performance and the creation of feeding currents.
F. nereidis may have moved through our ancient seas swimming in an inverted position — rare across arthropods and analogous to that observed in anostracans and some cladocerans. This highlight the importance of the carapace morphology in palaeo-ecological reconstructions and show that the arthropod carapace was already both a morphologically and functionally diverse character in the Cambrian period.
 |
| Bivalved Cambrian Arthropods / Alejandro Izquierdo-López |
Their phylogenetic analysis reveals a potential new group of mandibulate deposits and suspension feeders with homonomous legs and segments — some lacking certain mandibulate characters, such as antennae or mandibles — which may be related to an adaptation to this ecological niche and further illustrate a case of convergent evolution with some branchiopod taxa.
These results suggest that the bivalved carapace could have been a basal trait for all Mandibulata or may even have had an earlier origin if this and the bivalved carapace of the Isoxyiidae were found to be homologous.
Homologies between arthropod carapaces, bivalved or not, and structures such as radiodont shields, non-crustaceomorph univalved carapaces (e.g. Burgessia, Naraoia) and head shields (e.g. fuxianhuiids, habeliids, nauplius) are still quite poorly understood. We will need to find more examples to fully flesh out a comprehensive evolutionary analysis on this trait.
Besides, new data and morphological revisions on key bivalved arthropods could reshape the present phylogenetic analyses. Nonetheless, Cambrian bivalved arthropods certainly show a high ecological and taxonomic disparity, that is increasingly contributing to the understanding of the evolution of early arthropods and the Cambrian period as a whole.
Top Image by the talented Danielle Dufault @MesozoicMuse. Composite Bivalved Cambrian Arthropods by Alejandro Izquierdo-López.
Illustration: Interpretative cladogram based on a consensus tree from a Bayesian analysis using a Markov k model on a morphological dataset with 90 taxa and 213 characters. There is some interpretation here. Numbers next to nodes are posterior probabilities. The yellow box indicates the new monophyletic group to which Fibulacaris belongs. The green box highlights the group Hymenocarina.
Reference link: A Burgess Shale mandibulate arthropod with a pygidium: a case of convergent evolution. https://onlinelibrary.wiley.com/doi/abs/10.1002/spp2.1366